
 Thomas C. Südhof (Gotinga, Alemania; 22 de diciembre de 1955) es un neurobiólogo galardonado en el año 2013 con el premio Nobel de Medicina por sus estudios sobre el tráfico de vesículas en la célula. Südhof acaba de publicar un artículo en la revista Cell, una de las mejores del mundo, sobre las neurexinas, unas de las proteínas más interesantes en la investigación sobre el autismo.
Thomas C. Südhof (Gotinga, Alemania; 22 de diciembre de 1955) es un neurobiólogo galardonado en el año 2013 con el premio Nobel de Medicina por sus estudios sobre el tráfico de vesículas en la célula. Südhof acaba de publicar un artículo en la revista Cell, una de las mejores del mundo, sobre las neurexinas, unas de las proteínas más interesantes en la investigación sobre el autismo.
Las sinapsis son estructuras de unión y comunicación entre neuronas. Están formados por un elemento presináptico y un elemento postsináptico, partes de dos neuronas diferentes. Es decir, en la sinapsis la información salta de una neurona a otra y la segunda queda excitada, inhibida o igual, en función de la acción de la primera. Las sinapsis permiten la transmisión y computación de información, haciendo que las neuronas se conecten en billones de circuitos, redes que presentan numerosos solapamientos y que están intensamente interdigitadas unas con otras. Cómo de esa maraña increíble surgen pensamientos, sentimientos, memorias, es una de las cuestiones más asombrosas de la biología moderna.
Durante el desarrollo, las neuronas nacen, migran y extienden sus axones, a menudo a través de largas distancias hasta alcanzar sus dianas específicas. Entonces estos axones forman sinapsis sobre estas otras neuronas postsinápticas y van formando los circuitos neuronales. La formación de sinapsis tiene tres componentes básicos: reconocimiento de la célula diana, organización de los componentes básicos de la sinapsis tales como vesículas sinápticas y zonas activas y, finalmente, la especificación de las propiedades de la sinapsis, tales como identidad del transmisor, probabilidad de liberación de vesículas, composición de la zona receptora postsináptica o competencia para plasticidad a corto y largo plazo. La formación de sinapsis va seguida muy frecuentemente de la eliminación de la sinapsis lo que hace que haya una renovación constante y que se produzcan muchas más sinapsis de las que finalmente existen: en la adolescencia el 40-50% de las sinapsis son eliminadas. La formación de sinapsis, por otro lado, no muestra un único «producto terminado» sino que se genera un continuo de estados sinápticos desde un contacto inicial hasta distintos tipos de sinapsis maduras.
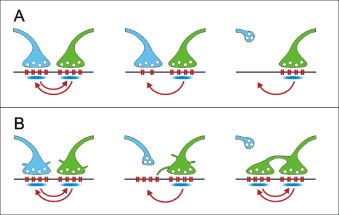 Es un patrón espacial curioso: la neurogénesis de neuronas de proyección y la formación de vías axonales a larga distancia son fenómenos restringidos a las etapas del desarrollo mientras que las sinapsis son formadas y eliminadas durante toda la vida. Las neuronas pierden entre un 5 y un 10% de las sinapsis cada mes dependiendo de la edad del animal y el tipo de neurona, pero algunas llegan a reemplazar a un 40% de las sinapsis cada mes (por ejemplo las sinapsis de las neuronas de capa VI sobre las capas I y II de la corteza sensorial del ratón). Las sinapsis recién formadas son menos estables que las sinapsis maduras y son aquellas las que se recambian normalmente. Aunque hay una cierta dependencia de la actividad, en realidad es un efecto muy bajo y la importancia de la actividad de un circuito sobre la formación y eliminación de sinapsis es sorprendentemente modesta. Por otro lado las sinapsis son asombrosamente plásticas pueden cambiar no solo su fuerza sino también, por ejemplo, el neurotransmisor que están liberando.
Es un patrón espacial curioso: la neurogénesis de neuronas de proyección y la formación de vías axonales a larga distancia son fenómenos restringidos a las etapas del desarrollo mientras que las sinapsis son formadas y eliminadas durante toda la vida. Las neuronas pierden entre un 5 y un 10% de las sinapsis cada mes dependiendo de la edad del animal y el tipo de neurona, pero algunas llegan a reemplazar a un 40% de las sinapsis cada mes (por ejemplo las sinapsis de las neuronas de capa VI sobre las capas I y II de la corteza sensorial del ratón). Las sinapsis recién formadas son menos estables que las sinapsis maduras y son aquellas las que se recambian normalmente. Aunque hay una cierta dependencia de la actividad, en realidad es un efecto muy bajo y la importancia de la actividad de un circuito sobre la formación y eliminación de sinapsis es sorprendentemente modesta. Por otro lado las sinapsis son asombrosamente plásticas pueden cambiar no solo su fuerza sino también, por ejemplo, el neurotransmisor que están liberando.
Hay dos características de los circuitos neuronales que son llamativas.  La primera es que el mapa de las conexiones sinápticas de un circuito determinado (el diagrama de cableado) no permite predecir las relaciones de entradas y salidas porque las sinapsis tienen propiedades plásticas y diversas. Es decir, las sinapsis no son unidades lineales de información y procesan de manera específica la información codificada por los patrones de disparo de la neurona presináptica. Esta transformación de los potenciales de acción de la neurona presináptica en las señales de la neurona postsináptica en los miles de sinapsis de entrada y de salida de una neurona varía dramáticamente entre diferentes sinapsis en función de sus propiedades y de su plasticidad y, además, es modulada por señales difusibles (por ejemplo, los endocannabinoides) y por entradas moduladores (p.e. la dopamina y la serotonina). El resultado es que una neurona integra señales múltiples en un patrón de descarga de potenciales de acción, que a su vez es transformado en una señal de salida que no solo varía entre distintas neuronas sino también entre los miles de sinapsis de una misma neurona.
La primera es que el mapa de las conexiones sinápticas de un circuito determinado (el diagrama de cableado) no permite predecir las relaciones de entradas y salidas porque las sinapsis tienen propiedades plásticas y diversas. Es decir, las sinapsis no son unidades lineales de información y procesan de manera específica la información codificada por los patrones de disparo de la neurona presináptica. Esta transformación de los potenciales de acción de la neurona presináptica en las señales de la neurona postsináptica en los miles de sinapsis de entrada y de salida de una neurona varía dramáticamente entre diferentes sinapsis en función de sus propiedades y de su plasticidad y, además, es modulada por señales difusibles (por ejemplo, los endocannabinoides) y por entradas moduladores (p.e. la dopamina y la serotonina). El resultado es que una neurona integra señales múltiples en un patrón de descarga de potenciales de acción, que a su vez es transformado en una señal de salida que no solo varía entre distintas neuronas sino también entre los miles de sinapsis de una misma neurona.
Lo segundo es que la mayoría de las neuronas del encéfalo participan simultáneamente en muchos circuitos diferentes, no solo en uno, y suele ser muy difícil identificar los distintos circuitos en los que participa una neurona determinada. El enfoque habitual para analizar circuitos es monitorizar la actividad de un tipo determinado de neuronas durante un comportamiento concreto, determinar los efectos sobre ese comportamiento de silenciar o activar estas neuronas y, en tercer lugar, cartografiar las conexiones sinápticas de las neuronas analizadas. Sin embargo, los datos que conseguimos con estas estrategias experimentales no nos permiten entender cómo funciona una sinapsis dentro de un circuito neuronal ni saber qué hacen otras neuronas del circuito y cómo contribuye cada sinapsis de cada neurona a ese comportamiento.
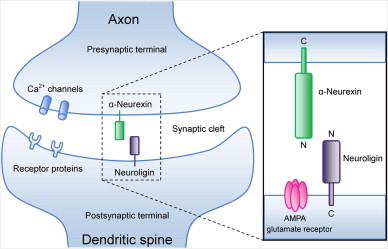 Südhof plantea que el establecimiento, propiedades y dinámica de las sinapsis están gobernadas por una lógica molecular que es controlada por distintas moléculas de señalización transsináptica; es decir, hay elementos pre y postsinápticos que se enlazan entre sí y eso determina dónde se forma una sinapsis, entre qué neuronas, cómo actúa esa sinapsis y qué resulta de su acción en la neurona postsináptica. Südhof se centra en esta revisión en las neurexinas por tres razones: son las moléculas de la sinapsis de las que hay más estudios en profundidad; son un elemento fundamental en distintos trastornos incluido el autismo y, tercero, pueden ser un ejemplo perfecto del conjunto de las interacciones sinápticas. Las neurexinas tienen un dominio extracellar que interacciona con otras proteínas en la hendidura sináptica, en particular con las neuroliginas. La neurexina y la neuroligina «se dan la mano» lo que establece la conexión entre las dos neuronas y la formación de una sinapsis.
Südhof plantea que el establecimiento, propiedades y dinámica de las sinapsis están gobernadas por una lógica molecular que es controlada por distintas moléculas de señalización transsináptica; es decir, hay elementos pre y postsinápticos que se enlazan entre sí y eso determina dónde se forma una sinapsis, entre qué neuronas, cómo actúa esa sinapsis y qué resulta de su acción en la neurona postsináptica. Südhof se centra en esta revisión en las neurexinas por tres razones: son las moléculas de la sinapsis de las que hay más estudios en profundidad; son un elemento fundamental en distintos trastornos incluido el autismo y, tercero, pueden ser un ejemplo perfecto del conjunto de las interacciones sinápticas. Las neurexinas tienen un dominio extracellar que interacciona con otras proteínas en la hendidura sináptica, en particular con las neuroliginas. La neurexina y la neuroligina «se dan la mano» lo que establece la conexión entre las dos neuronas y la formación de una sinapsis.
Las neurexinas son proteínas muy conservadas durante la evolución que se expresan en miles de isoformas diferente por un splicing alternativo. Es decir, en condiciones «básicas», un gen de ADN, genera un ARN mensajero, que se traduce a una proteína. Pero para algunos genes, incluidos los tres que en mamíferos codifican las neurexinas, el ARN mensajero es procesado de muchas maneras diferentes, se le hacen distintos cortes que eliminan parte de la secuencia y el resultado es que las proteínas que se forman con esos ARNm alternativos son distintas, lo que hemos denominado isoformas. La enorme variedad de neurexinas es un componente central de ese código dinámico de las sinapsis. Además, las neurexinas son presinápticas y se pueden unir a distintos elementos postsinápticos, regulando de este modo las propiedades de la sinapsis. Entre los distintos tipos de ligandos postsinápticos están las neuroliginas, los complejos cerebelina/GluD y las latrofilinas. Estas uniones modulan la relación entre las entradas y las salidas de los circuitos neuronales de los que forman parte.
Es decir, en condiciones «básicas», un gen de ADN, genera un ARN mensajero, que se traduce a una proteína. Pero para algunos genes, incluidos los tres que en mamíferos codifican las neurexinas, el ARN mensajero es procesado de muchas maneras diferentes, se le hacen distintos cortes que eliminan parte de la secuencia y el resultado es que las proteínas que se forman con esos ARNm alternativos son distintas, lo que hemos denominado isoformas. La enorme variedad de neurexinas es un componente central de ese código dinámico de las sinapsis. Además, las neurexinas son presinápticas y se pueden unir a distintos elementos postsinápticos, regulando de este modo las propiedades de la sinapsis. Entre los distintos tipos de ligandos postsinápticos están las neuroliginas, los complejos cerebelina/GluD y las latrofilinas. Estas uniones modulan la relación entre las entradas y las salidas de los circuitos neuronales de los que forman parte.
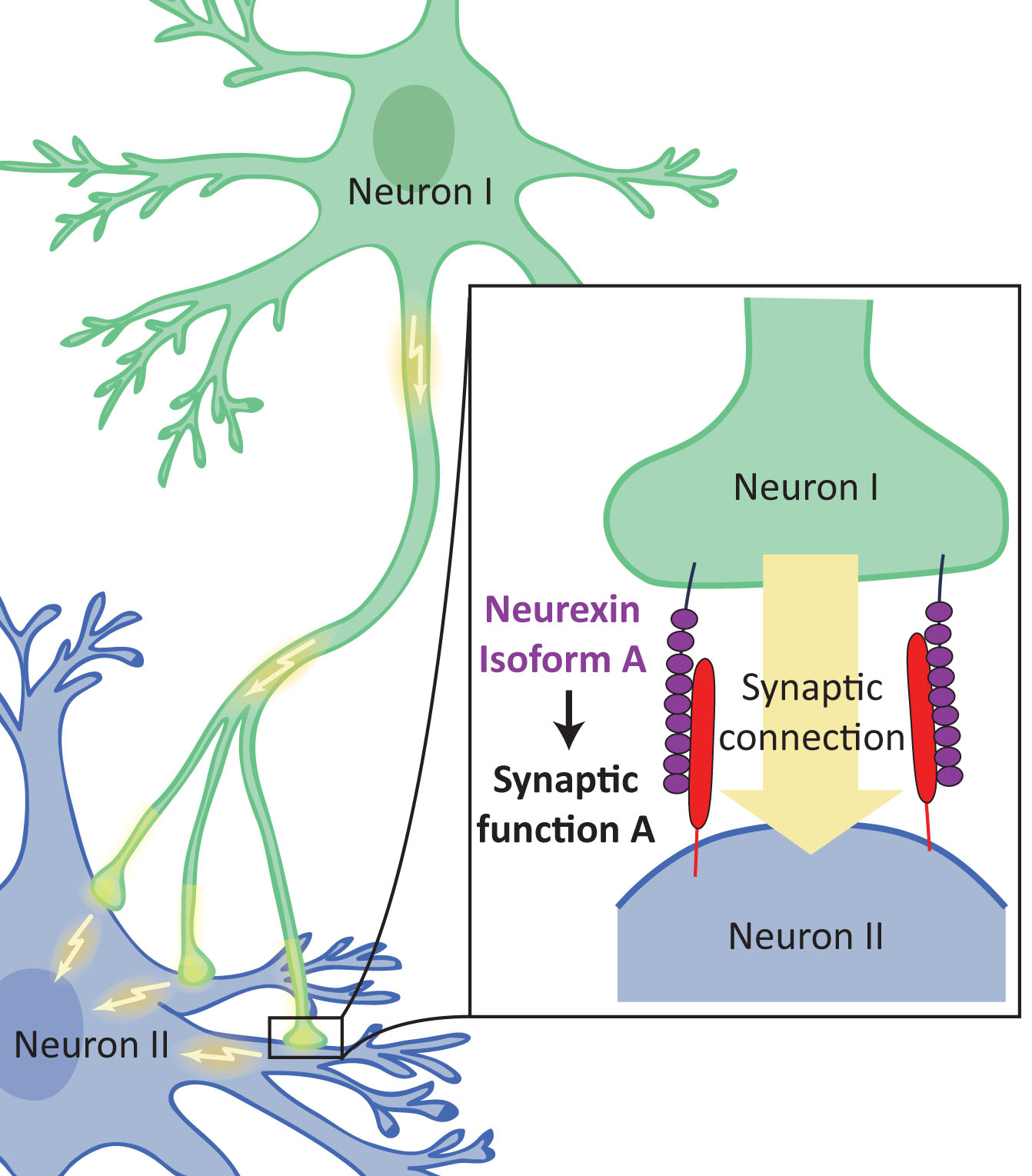 Las neurexinas son el núcleo de una red de señalización transsináptica que controla las propiedades de la sinapsis y de esta manera determinan las respuestas exactas de las sinapsis a los patrones de potenciales de acción de una neurona y de un circuito y que además son vulnerables y tienen que ver con la aparición de un autismo. En 2007 se hizo un estudio del ADN de 1168 familias donde había al menos un caso de autismo y se encontró la implicación de una región del cromosoma 11 donde está el gen neurexin I, que tiene un papel activo durante el desarrollo temprano del cerebro.
Las neurexinas son el núcleo de una red de señalización transsináptica que controla las propiedades de la sinapsis y de esta manera determinan las respuestas exactas de las sinapsis a los patrones de potenciales de acción de una neurona y de un circuito y que además son vulnerables y tienen que ver con la aparición de un autismo. En 2007 se hizo un estudio del ADN de 1168 familias donde había al menos un caso de autismo y se encontró la implicación de una región del cromosoma 11 donde está el gen neurexin I, que tiene un papel activo durante el desarrollo temprano del cerebro.
Los avances recientes en micromatrices y secuenciación de ADN, combinados con la colaboración de pacientes y sus familias, han permitido una mirada como nunca antes se había logrado sobre la base genética y molecular de los trastornos neurales. Estos estudios han generado tres descubrimientos sorprendentes: primero que el genoma humano es llamativamente fluido. Un ejemplo es la presencia de delecciones y duplicaciones del ADN, que generan las llamadas variaciones en el número de copias (VNC) que se producen de forma habitual en cada nueva generación, produciendo una dinámica genética que nadie esperaba. En segundo lugar, mutaciones en muchos genes diferentes predisponen para muchos trastornos neurales, probablemente cientos de genes en los casos del autismo y la esquizofrenia. Tercero, para muchas mutaciones, las presentaciones clínicas varían mucho de una persona afectada a otra a pesar de tener una mutación similar.  Hay también problemas: a menudo se encuentra que una mutación va asociada a una enfermedad pero se le da menos importancia cuando se ve que también está asociada a otros trastornos cuando debería considerarse una señal de que puede haber algún paso común afectado y ese gen debería recibir más y no menos interés. Otro problema es que muchos estudios abordan un trastorno con una técnica, por ejemplo un análisis con micromatrices para detectar VNC o una secuenciación del exoma para detectar mutaciones puntuales en vez de intentar barrer sistemáticamente el genoma de una población de pacientes con un abordaje experimental que use múltiples técnicas. Eso evitaría la situación en la que nos encontramos donde es a menudo difícil comparar los resultados de unos estudios con otros. Finalmente en muchos de los genes identificados no se conoce su función con lo que nos encontramos en la situación de intentar evaluar la importancia de las mutaciones y las relaciones entre los genes afectados sin tener datos directos. Quizá el camino es aceptar que la lista de genes es solo un primer paso y que las discusiones y elucubraciones sobre su importancia deben postponerse hasta que tengamos una imagen más completa del paisaje genético de un trastorno como el autismo.
Hay también problemas: a menudo se encuentra que una mutación va asociada a una enfermedad pero se le da menos importancia cuando se ve que también está asociada a otros trastornos cuando debería considerarse una señal de que puede haber algún paso común afectado y ese gen debería recibir más y no menos interés. Otro problema es que muchos estudios abordan un trastorno con una técnica, por ejemplo un análisis con micromatrices para detectar VNC o una secuenciación del exoma para detectar mutaciones puntuales en vez de intentar barrer sistemáticamente el genoma de una población de pacientes con un abordaje experimental que use múltiples técnicas. Eso evitaría la situación en la que nos encontramos donde es a menudo difícil comparar los resultados de unos estudios con otros. Finalmente en muchos de los genes identificados no se conoce su función con lo que nos encontramos en la situación de intentar evaluar la importancia de las mutaciones y las relaciones entre los genes afectados sin tener datos directos. Quizá el camino es aceptar que la lista de genes es solo un primer paso y que las discusiones y elucubraciones sobre su importancia deben postponerse hasta que tengamos una imagen más completa del paisaje genético de un trastorno como el autismo.
Para leer más:
- Anderson GR, Aoto J, Tabuchi K, Földy C, Covy J, Yee AX, Wu D, Lee SJ, Chen L, Malenka RC, Südhof TC (2015) β-Neurexins Control Neural Circuits by Regulating Synaptic Endocannabinoid Signaling. Cell 62(3): 593-606.
- Südhof TC (2017) Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell 171(4): 745-769.

Responder a Victoria LeyCancelar respuesta