
 En 1846, Rudolf Virchow observó unas células en las paredes de los ventrículos cerebrales y postuló en uno de los libros científicos más famosos del siglo XIX, su Patología Celular (1858), que el cerebro contenía una segunda población de células, distintas a las neuronas. Pensó que eran parte de un Bindegewebe, un tejido conjuntivo, y las llamó glía o Nervenkitt. Normalmente se traduce como «pegamento nervioso» o «cemento nervioso» pero el idioma alemán tiene otras palabras para eso, tales como «Leim», «Klebstoff» y «Zement». Al usar la palabra Kitt se refería a algo pegajoso pero que tenía volumen y forma, «un tipo de masilla en el que están inmersos los elementos neuronales», algo cuyo equivalente podría ser la plastilina o la masilla de cristalero, una pasta que mantenía unidas a las neuronas. Plantear que eran tejido conjuntivo establecía una analogía con el sistema nervioso periférico donde los nervios tienen una envuelta de conjuntivo que rodea a los axones.
En 1846, Rudolf Virchow observó unas células en las paredes de los ventrículos cerebrales y postuló en uno de los libros científicos más famosos del siglo XIX, su Patología Celular (1858), que el cerebro contenía una segunda población de células, distintas a las neuronas. Pensó que eran parte de un Bindegewebe, un tejido conjuntivo, y las llamó glía o Nervenkitt. Normalmente se traduce como «pegamento nervioso» o «cemento nervioso» pero el idioma alemán tiene otras palabras para eso, tales como «Leim», «Klebstoff» y «Zement». Al usar la palabra Kitt se refería a algo pegajoso pero que tenía volumen y forma, «un tipo de masilla en el que están inmersos los elementos neuronales», algo cuyo equivalente podría ser la plastilina o la masilla de cristalero, una pasta que mantenía unidas a las neuronas. Plantear que eran tejido conjuntivo establecía una analogía con el sistema nervioso periférico donde los nervios tienen una envuelta de conjuntivo que rodea a los axones.  Más aún, Virchow era un gran patólogo y estaba familiarizado con los procesos inflamatorios que afectaban a las cavidades ventriculares. Para él, solo el tejido conjuntivo podía inflamarse; por lo tanto, bajo el epitelio que bordeaba los ventrículos debía haber algo de conjuntivo.
Más aún, Virchow era un gran patólogo y estaba familiarizado con los procesos inflamatorios que afectaban a las cavidades ventriculares. Para él, solo el tejido conjuntivo podía inflamarse; por lo tanto, bajo el epitelio que bordeaba los ventrículos debía haber algo de conjuntivo.
Otros investigadores habían visto células gliales anteriormente pero ninguno las había puesto nombre y no estaba claro qué eran o si no eran más que un tipo especial de neuronas. El primero que documentó células gliales fue el fisiólogo francés Henri Dutrochet en 1824. No tenía ni idea de qué era aquello y simplemente anotó que había glóbulos entre los nervios de los moluscos, un lugar donde ahora sabemos que no hay neuronas. Otros pioneros postularon cosas sobre la glía que se demostrarían ciertas un siglo más tarde: su capacidad para secretar sustancias químicas (Nageotte), su asociación con los vasos sanguíneos (Golgi), su plasticidad morfológica (Cajal), su capacidad para actuar como un aislante (Cajal), su papel en la captación de neurotransmisor y en la finalización de la transmisión (Lugaro) y su implicación en algunas enfermedades (Virchow). Para Cajal, el pionero en la identificación en el sistema nervioso central de células que no eran neuronas fue Otto Deiters.  Deiters había razonado correctamente que si una célula no tenía axón («Hauptaxencylinderfortsatz» o «prolongación cilindro eje principal») no podía ser una neurona. En su libro póstumo de 1865 aparecen dos células marcadas como Bindegewebszellen, células del tejido conjuntivo y, de hecho, durante un tiempo la glía se conoció como las células de Deiters.
Deiters había razonado correctamente que si una célula no tenía axón («Hauptaxencylinderfortsatz» o «prolongación cilindro eje principal») no podía ser una neurona. En su libro póstumo de 1865 aparecen dos células marcadas como Bindegewebszellen, células del tejido conjuntivo y, de hecho, durante un tiempo la glía se conoció como las células de Deiters.
Otros investigadores fueron identificando distintos tipos celulares en el tejido nervioso que ahora incluimos dentro de la glía. Theodor Schwann vio unas células en el sistema nervioso periférico y ahora las llamamos células de Schwann y sabemos que se encargan de formar la envuelta mielínica de los nervios. De forma similar, Heinrich Müller, un profesor de Würzburg, observó en la retina unas células radiales y ahora las llamamos las células de Müller. Los primeros que describieron una malla de células gliales en la corteza cerebral fueron Friedrich Merkel y Jacob Henle, mientras que Mihály von Lenhossék, un húngaro que trabajaba en Alemania, las denominó astrocitos por su forma estrellada. La primera imagen convincente de un oligodendrocito fue publicada por Camillo Golgi quien dijo que la característica definida por Deiters, la ausencia de un axón, era el único rasgo fiable. Lenhossék cita a Golgi como el primero que vio fibras extendiéndose rápidamente desde las células ependimarias hasta la superficie del cerebro pero dice que fue Fridtjof Nansen, el neurocientífico que se convirtió en explorador polar, el primero que postuló que derivaban del epéndimo embrionario.  Esto era algo importante, porque el epéndimo es ectodérmico, es decir similar a los epitelios y al tejido nervioso y por lo tanto diferente del tejido conjuntivo. La glía entonces era mucho más próxima a las neuronas, eran por así decirlo parientes cercanos, mientras que el conjuntivo estaba alejado ontogenética y evolutivamente. El origen ependimario de las células gliales fue confirmado por Ramón y Cajal y su discípulo, Nicolás Achúcarro habló de «glía ependimaria» y «glía autónoma», las demás células, para reforzar la relación entre ambos tipos. En 1889, Wilhelm His dio otro espaldarazo a la importancia de las células gliales en el sistema nervioso al postular que las largas prolongaciones de la glía radial servían como sistemas de guía para la migración de las neuronas en desarrollo.
Esto era algo importante, porque el epéndimo es ectodérmico, es decir similar a los epitelios y al tejido nervioso y por lo tanto diferente del tejido conjuntivo. La glía entonces era mucho más próxima a las neuronas, eran por así decirlo parientes cercanos, mientras que el conjuntivo estaba alejado ontogenética y evolutivamente. El origen ependimario de las células gliales fue confirmado por Ramón y Cajal y su discípulo, Nicolás Achúcarro habló de «glía ependimaria» y «glía autónoma», las demás células, para reforzar la relación entre ambos tipos. En 1889, Wilhelm His dio otro espaldarazo a la importancia de las células gliales en el sistema nervioso al postular que las largas prolongaciones de la glía radial servían como sistemas de guía para la migración de las neuronas en desarrollo.
Sin embargo, la cosa distaba de estar aclarada por la diversidad de las células gliales. Franz Nissl había visto unas células en la década de 1880 que se parecían a los macrófagos. Victor Babeş, estudiando un caso de rabia en 1897 vio que esas células se activaban y mostraban ramificaciones, algo que se observaba en otras enfermedades infecciosas. En 1913 Cajal planteaba que además de las dos categorías principales: neuronas y neuroglia «legítima» existía un «tercer elemento», células sin prolongaciones que parecían derivar del mesodermo. Pío del Río-Hortega demostró a su maestro que eran células incompletamente teñidas y que correspondían en realidad a oligodendrocitos, un tipo definido por él en 1922, o a un nuevo tipo, al que llamó microglía y que durante décadas fueron conocidas como las células de Hortega.
Don Pío, el mejor discípulo de Cajal y candidato al premio Nobel en 1929 y 1937, fue el que puso orden en todo este caos, confirmando que la glía estaba formada por células individuales y clasificando los distintos grupos.  Comparó las células oligodendrogliales con las células de Schwann, llegando a la acertado conclusión que las dos estaban relacionadas con las envueltas de mielina, unas estructuras que mostraban alteraciones en algunas enfermedades como la esclerosis múltiple. Cajal diferenció dos tipos de astrocitos: fibrosos y protoplasmáticos y Río-Hortega demostró la respuesta de la microglía al daño cerebral planteando que la función de la microglía era la fagocitosis de elementos dañinos o restos celulares, indicando que «si hay neuronofagia, creemos que solo las células de la microglía lo hacen, ya que su capacidad para migrar y fagocitar está fuera de toda duda», una afirmación que se anticipaba a la investigación sobre la microglía publicada décadas después. En 1988 William F. Hickey e Hiromitsu Kimura publicaron en Science que las células microgliales perivasculares derivaban de la médula ósea y expresaban proteínas del complejo mayor de histocompatibilidad (clase II), lo que confirmaba los postulados de Río Hortega de que la microglía funcionaba de forma similar a los macrófagos haciendo fagocitosis y presentación de antígenos. Las distintas partes del puzle iban siendo identificadas.
Comparó las células oligodendrogliales con las células de Schwann, llegando a la acertado conclusión que las dos estaban relacionadas con las envueltas de mielina, unas estructuras que mostraban alteraciones en algunas enfermedades como la esclerosis múltiple. Cajal diferenció dos tipos de astrocitos: fibrosos y protoplasmáticos y Río-Hortega demostró la respuesta de la microglía al daño cerebral planteando que la función de la microglía era la fagocitosis de elementos dañinos o restos celulares, indicando que «si hay neuronofagia, creemos que solo las células de la microglía lo hacen, ya que su capacidad para migrar y fagocitar está fuera de toda duda», una afirmación que se anticipaba a la investigación sobre la microglía publicada décadas después. En 1988 William F. Hickey e Hiromitsu Kimura publicaron en Science que las células microgliales perivasculares derivaban de la médula ósea y expresaban proteínas del complejo mayor de histocompatibilidad (clase II), lo que confirmaba los postulados de Río Hortega de que la microglía funcionaba de forma similar a los macrófagos haciendo fagocitosis y presentación de antígenos. Las distintas partes del puzle iban siendo identificadas.
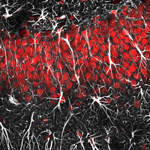
La glía son células algo más pequeñas que las neuronas, en un número parecido (85.000 millones de células gliales frente a 86.000 millones de neuronas en el cerebro humano) y que ocupan aproximadamente la mitad del volumen del encéfalo y de la médula espinal, aunque la proporción neuronas:glía varía de una zona a otra. En la actualidad se distinguen dentro de la glía dos grandes grupos: microglía y macroglía, diferenciándose dentro de esta cuatro tipos especializados: células ependimarias, células de Schwann, oligodendrocitos y astroglía.
 La astroglía incluye los astrocitos, la glía marginal, la glía radial, las células de Bergmann en el cerebelo, las células de Müller en la retina, los pituicitos en la neurohipófisis y los tanicitos en el hipotálamo. En la corteza cerebral, la glía más común son los oligodendrocitos (75,6%) seguidos por los astrocitos (17,3%) y la más escasa es la microglía (6,5%).
La astroglía incluye los astrocitos, la glía marginal, la glía radial, las células de Bergmann en el cerebelo, las células de Müller en la retina, los pituicitos en la neurohipófisis y los tanicitos en el hipotálamo. En la corteza cerebral, la glía más común son los oligodendrocitos (75,6%) seguidos por los astrocitos (17,3%) y la más escasa es la microglía (6,5%).
En 1909 Ramón y Cajal se preguntó «¿cuál es la función de la glía?» y él mismo dio la respuesta «Nadie lo sabe». Virchow pensaba que lo que hacían era mantener unidas a las células y dar forma al cerebro pero pronto estuvo claro que hacían muchas más cosas. Golgi decía que alimentaban a las neuronas pero él pensaba equivocadamente que las dendritas servían para conseguir nutrientes y no intervenían en la transmisión nerviosa. Para ello se basaba en la semejanza entre las prolongaciones de la glía y las dendritas de las neuronas y que algunas de esas prolongaciones se acercaban hasta los vasos sanguíneos a los que rodeaban, los llamados pies chupadores. Cajal rechazó las ideas de Golgi y propuso que los procesos de la glía podían servir para aislar los axones, no solo por la formación de las envueltas de mielina sino también por situarse entre los axones amielínicos. También se vio que las células gliales modificaban su morfología y el joven Wilder Penfield, trabajando con Río-Hortega en Madrid, confirmó que las células en bastoncito (Stäbchenzellen) era microglía en movimiento.
Ya en ese inicio del siglo XX, la noción de Virchow de que la glía era una masilla que simplemente mantenía unidas a las neuronas se fue resquebrajando. Desde entonces, y ya ha pasado más de un siglo, los neurocientíficos fueron descubriendo que eran mucho más activas e importantes.  Durante gran parte del siglo el papel de las células gliales era bastante pasivo: establecían el andamiaje del cerebro, los nutrientes pasaban a su través y se encargaban de formar las envueltas de mielina y de hacer la limpieza. Es decir, la glía eran los arquitectos del cerebro, los médicos, la policía y los barrenderos y jardineros pero en las dos últimas décadas, el interés por la glía se disparó y nuevas funciones se fueron sumando. Hacían cosas sorprendentes entre las que podemos citar las siguientes:
Durante gran parte del siglo el papel de las células gliales era bastante pasivo: establecían el andamiaje del cerebro, los nutrientes pasaban a su través y se encargaban de formar las envueltas de mielina y de hacer la limpieza. Es decir, la glía eran los arquitectos del cerebro, los médicos, la policía y los barrenderos y jardineros pero en las dos últimas décadas, el interés por la glía se disparó y nuevas funciones se fueron sumando. Hacían cosas sorprendentes entre las que podemos citar las siguientes:
La microglía es el principal sistema de retirada de basura del cerebro pero no solo eso. Estas células gliales se encargan de eliminar el exceso de fluido, retirar y digerir las células muertas y eliminar cualquier resto celular que pueda alterar el funcionamiento normal del cerebro. La microglía patrulla por el cerebro pero además elimina acúmulos de beta amiloide y otras proteínas asociadas con la enfermedad de Alzheimer. La microglía también interviene en el podado de conexiones sinápticas que se producen en el desarrollo cerebral.
Las células precursoras de los oligodendrocitos (CPO) son especialmente activas. Con el tiempo pueden madurar y convertirse en oligodendrocitos que envuelven los axones formando las cubiertas aislantes de mielina. Un único oligodendrocito envuelve múltiples axones, cada uno de una neurona.  Pero las CPO establecen sinapsis con las neuronas y cambian su propio comportamiento en función de las señales eléctricas que reciben de ellas. Hay evidencias de que la comunicación entre CPO y oligodendrocitos va en los dos sentidos y alcanza a otras neuronas. Por poner un ejemplo, la superficie de los CPO está recubierta de una proteína llamada NG2. Los impulsos eléctricos que las CPO reciben de las neuronas ponen en marcha enzimas que despegan la NG2 de la membrana plasmática haciendo que la proteína fluya y contacte con otras neuronas cercanas, un nuevo tipo de comunicación intercelular. Al llegar a esas otras neuronas las hace más sensibles a transmisores como el glutamato, uno de los principales mensajeros intercelulares. Si el proceso se bloquea eliminando NG2 o sus enzimas, los ratones se vuelven más lentos y menos curiosos. También se ha visto que los oligodendrocitos y las neuronas están acoplados metabólicamente, intercambiando materiales como lactato que permite obtener energía. Además los oligodendrocitos producen factores neuromoduladores que ajustan la comunicación entre las neuronas.
Pero las CPO establecen sinapsis con las neuronas y cambian su propio comportamiento en función de las señales eléctricas que reciben de ellas. Hay evidencias de que la comunicación entre CPO y oligodendrocitos va en los dos sentidos y alcanza a otras neuronas. Por poner un ejemplo, la superficie de los CPO está recubierta de una proteína llamada NG2. Los impulsos eléctricos que las CPO reciben de las neuronas ponen en marcha enzimas que despegan la NG2 de la membrana plasmática haciendo que la proteína fluya y contacte con otras neuronas cercanas, un nuevo tipo de comunicación intercelular. Al llegar a esas otras neuronas las hace más sensibles a transmisores como el glutamato, uno de los principales mensajeros intercelulares. Si el proceso se bloquea eliminando NG2 o sus enzimas, los ratones se vuelven más lentos y menos curiosos. También se ha visto que los oligodendrocitos y las neuronas están acoplados metabólicamente, intercambiando materiales como lactato que permite obtener energía. Además los oligodendrocitos producen factores neuromoduladores que ajustan la comunicación entre las neuronas.
Otro tipo glial, los astrocitos, han demostrado ser los más complejos, interviniendo en funciones muy diversas:
- Unidad neurovascular. Los astrocitos envuelven perfectamente los vasos sanguíneos haciendo que todo lo que salga de la sangre hacia el tejido nervioso tenga que pasar a través de ellos. Así, si llega una sustancia tóxica la pararán como si fuesen los probadores de la comida de un rey, evitando que haga daño a las neuronas y asegurando una correcta homeostasis en el cerebro. También modulan el propio flujo sanguíneo en función de los niveles de oxígeno y CO2. La glía es imprescindible hasta para respirar. Si tenemos poco oxígeno y mucho CO2, baja el pH en la sangre. Los astrocitos del bulbo raquídeo, la parte del tronco encefálico que controla la respiración y el latido cardíaco, incrementan su señalización intracelular de calcio y empiezan a secretar ATP a su alrededor, una molécula que no solo almacena energía sino que hace muchas otras cosas en las células. Este ATP estimula a las neuronas a su alrededor a disparar, lo que incrementa la respiración, lo que lleva más oxígeno al cerebro. Si el pH sube en vez de bajar, el ritmo de respiración se hace más lento.

- Acoplamiento metabólico. Los astrocitos tienen transportadores de glucosa y son las únicas células capaces de almacenar energía en el cerebro adulto, en forma de glucógeno. Estos depósitos energéticos se movilizan mediante señales neuroactivas como la noradrenalina y el péptido intestinal vasoactivo. Tras las señales adecuadas, los astrocitos liberan lactato que es aprovechado por las neuronas.
- Participación en la plasticidad. Los astrocitos regulan la plasticidad sináptica y se encargan de la eliminación de sinapsis inoperantes o superfluas y de la formación de nuevos contactos. Algunas células gliales excretan moléculas que promueven la formación de nuevas conexiones entre las neuronas mientras que otros rodean y digieren las sinapsis que están siendo poco utilizadas, cambiando las conexiones cerebrales todos los días, durante toda la vida. Participan también en funciones cognitivas superiores como el aprendizaje y la memoria. Tras el aprendizaje, se produce un incremento (upregulation) de los genes implicados en el acoplamiento entre astrocitos y neuronas.
- Gliotransmisión. Los astrocitos presentan numerosos receptores y canales iónicos similares a los de las neuronas. Pueden también liberar neurotransmisores, eliminarlos de la sinapsis y controlar cuanto transmisor liberará una neurona en el futuro. En muchas sinapsis glutamatérgicas, los astrocitos envuelven la hendidura sináptica y comunican con el elemento presináptico y el postsináptico, en lo que se ha llamado una sinapsis tripartita.
- Función neurotrófica. Los astrocitos fabrican y liberan muchos factores tróficos como el FGF2, el BDNF, necesarios para la supervivencia de las neuronas, y otros factores de crecimiento que modulan la transmisión sináptica y la plasticidad. Esto tiene efecto en la cognición y en los comportamientos asociados al estado de ánimo.
- Respuesta a la lesión. Tras un daño cerebral, los astrocitos se convierten en reactivos, modifican su morfología y cambian su expresión de genes. Pueden luchar también contra patógenos modificando la permeabilidad de la barrera hematoencefálica y secretar citoquinas que atraen a células inmunitarias desde la circulación sanguínea para luchar contra el daño cerebral.

- También intervienen en la modulación y la recaptación de los neurotransmisores y parcelan la sustancia gris en compartimentos funcionales cuyos niveles umbral de activación y las eferencias hacia el exterior son definidas por las células gliales. La red de microdominios delimitada por los astrocitos regula la actividad neuronal y el flujo sanguíneo generando una coordinación marcada por la glía. Ahora sabemos que las células gliales son elementos muy dinámicos, enormemente versátiles e imprescindibles para que las neuronas realicen su función. La idea en la actualidad es que las células gliales son las compañeras imprescindibles de las neuronas.
En resumen, la interpretación de la glía ha cambiado radicalmente en la última década y es posible que los resultados más sorprendentes se hayan encontrando en estas células. Estos estudios demuestran que la comunicación entre neuronas y su plasticidad está continuamente bajo la influencia de las células gliales.
Para leer más:
- Fink J (2016) Beyond the neuron: Emerging roles of glial cells in neuroscience. PLOS Neuro http://blogs.plos.org/neuro/2016/01/25/beyond-the-neuron-emerging-roles-of-glial-cells-in-neuroscience/#.VqZ4_OHate8.twitter
- Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, Harris TH, Kipnis J (2015) Structural and functional features of central nervous system lymphatic vessels. Nature 523(7560): 337-341.
- Nedergaard M, Ransom B, Goldman SA (2003) New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci 26(10): 523-530.
- Somjen GG (1988) Nervenkitt: notes on the history of the concept of neuroglia. Glia 1: 2-9.

Muchas gracias por comentar